

MOP (IHC-grade), µ-Opioid Receptor Antibody

Citations
KO-Validated





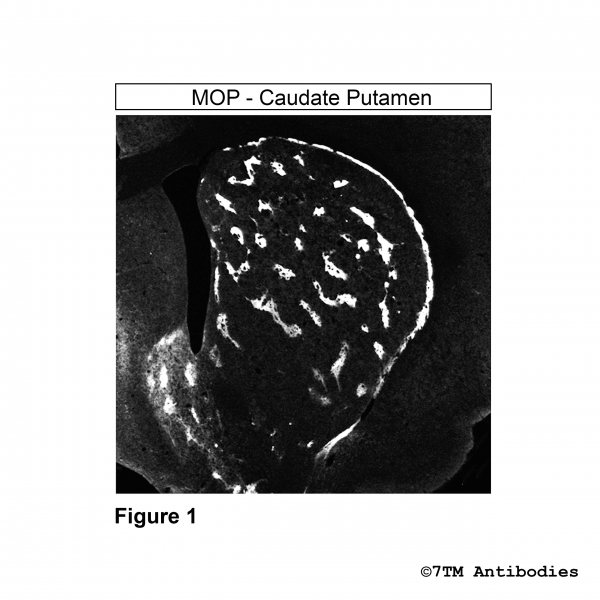




- Order number: 7TM0319N-IC
- Content: 100 µl
- Host: Rabbit




Citations

Citations

Citations

Citations

Citations
KO-Validated

Citations
KO-Validated

Citations

Citations
KO-Validated

Citations

Citations
KO-Validated

Citations

Citations

Citations

Citations
KO-Validated
, µ-Opioid Receptor Antibody")
Recently viewed